


Con el apoyo de CONARE, diferentes personas investigadoras de las universidades públicas llevaron a cabo una importante investigación para conocer más sobre la vulnerabilidad de algunos ecosistemas que nos sustentan. Analizando el estado actual de la delicada relación planta-polinizador en algunos páramos de Costa Rica.
En esta investigación se vigiló el equilibrio planta-polinizador en 3 páramos del Cerro de la Muerte, el principal macizo del norte de la Cordillera de Talamanca. En específico se localizó el área de investigación en:
- Reserva Biológica Cerro Las Vueltas
- Cerro Buena Vista (vertiente Pacífica)
- Cerros Sákira (vertiente del Caribe)
(Para ver los resultados de la investigación haga click aquí)
Un páramo consiste en una formación vegetal herbácea-arbustiva en donde predominan bambúes enanos, gramíneas, plantas en forma de roseta y arbustos, además de árboles generalmente esparcidos en algunas áreas (Stern & Kernan 2011). Estos páramos al igual que la mayoría de los ecosistemas terrestres, tienen gran dependencia de los polinizadores para mantener su equilibrio. Vale resaltar, que en América solo existen páramos en la Cordillera de Talamanca, en la Cordillera de los Andes y en el Caribe de Colombia.
Para resolver este proyecto los y las investigadoras definieron 4 objetivos. A continuación explicaremos dichos objetivos y cómo fueron resueltos.
Objetivo 1: Determinar el grado de especialización de los polinizadores a nivel de especie y de comunidad en redes de interacción planta-polinizador.
Este objetivo se cumplió por medio de la resolución de estas 4 metas:
1. Se identificarán 3 sitios que sean representativos del ecosistema de páramo.
2. Se cuantificarán las interacciones planta-polinizador en cada sitio de estudio.
3. Se identificarán insectos y plantas en el laboratorio que no hayan podido ser identificados en el campo.
4. Se calcularán índices de especialización ecológica a nivel de comunidad y de especies en redes planta-polinizador.
Objetivo 2: Analizar los patrones de variación temporal en abundancia de recursos florales y poblaciones de polinizadores en el ecosistema de páramo
Este objetivo se dividió en 3 metas para cumplirse, sin embargo aún está en desarrollo el análisis sobre variación en la abundancia de recursos florales. También debido a la gran variedad de insectos y las similitudes que presentan es necesario la colaboración de especialistas en el tema para llegar a una conclusión que tenga el margen de error más pequeño posible con respecto a las especies observadas.
1. Se calculará la abundancia de flores por especie de planta.
2. Se calculará la abundancia de los polinizadores.
– Se realizaron conteos de flores por especie de planta en transectos
– Se capturaron y anillaron colibríes
– Los análisis sobre variación en la abundancia de recursos florales están pendientes y serán incluidos en un artículo que está en proceso de elaboración.
3. Se identificarán insectos y plantas en el laboratorio que no hayan podido ser identificados en el campo.
Objetivo 3: Describir la estructura funcional de la comunidad de polinizadores e identificar rasgos funcionales asociados con respuestas potenciales de las especies de polinizadores al cambio climático.
Para este objetivo se concluyó la recolección de datos e información necesaria para el análisis funcional y la identificación de rasgos relevantes en organismos polinizadores, el cual sigue en desarrollo.
1. Se obtendrán mediciones de rasgos funcionales de los polinizadores.
2. Se analizará la diversidad funcional de la comunidad de polinizadores.
3. Se identificarán rasgos de respuesta de los polinizadores al cambio climático.
Objetivo 4: Analizar la capacidad de las especies de colibríes que habitan el páramo para utilizar nuevos recursos florales.
1. Se establecerán 3 sitios de parcelas experimentales en páramos.
2. Se definirán 3 especies de plantas con síndrome de ornitofilia que difieran en su morfología floral y se obtendrán mediciones de rasgos.
3. Se determinará la capacidad de supervivencia de especies de plantas a ser utilizadas en experimentos de trasplante.
4. Se determinará la capacidad de colibríes para utilizar flores de individuos trasplantados.
Resultados
Objetivo 1:
- Interacciones planta-colibrí
- Se observaron 3120 plantas de 49 especies diferentes. En total se registraron 892 interacciones entre 27 especies de plantas y 4 especies de colibríes.
- La proporción de especies de plantas que recibieron visitas de colibríes con respecto al total observadas fue del 55%. Las especies de colibríes que fueron observadas interactuando con plantas del páramo son Selasphorus flammula, Panterpe insignis, Colibri cyanotus y Eugenes fulgens los cuales visitaron un total de 340 flores.

Colibri cyanotus 
Eugenes fulgens 
Panterpe insignis 
Selasphorus flammula
- P. insignis es la única especie que ha sido reportada por ciertos estudios como residente común del Cerro de la Muerte, mientras que las otras especies son migrantes altitudinales que descienden en cierta época del año para pasar la época no reproductora en hábitats a menor elevación (Slud 1965, Wolf & Stiles 1970, Hainsworth & Wolf 1972)
- Durante las observaciones de campo realizadas a lo largo del proyecto se ha observado también a S. flammula como un habitante del páramo a lo largo de todo el año siendo los machos los más abundantes.
- Solo en una de las locaciones observadas hubo presencia de las 4 especies de colibrí analizadas. Esto fue en la Reserva Biológica Cerro Las Vueltas.
Especies de plantas visitadas por colibríes en tres sitios de páramo en el Cerro de la Muerte
| Orden | Familia | Especie |
| Asterales | Asteraceae | Pentacalia firmipes |
| Asterales | Campanulaceae | Centropogon valerioi |
| Celastrales | Celastraceae | Maytenus woodsonii |
| Ericales | Clethraceae | Clethra gelida |
| Ericales | Ericaceae | Cavendishia confertiflora |
| Ericales | Ericaceae | Comarostaphylis arbutoides |
| Ericales | Ericaceae | Macleania rupestris |
| Ericales | Ericaceae | Vaccinium consanguineum |
| Escalloniales | Escalloniaceae | Escallonia myrtilloides |
| Fabales | Polygalaceae | Monnina sylvatica |
| Gentianales | Gentianaceae | Halenia aquilegiella |
| Gentianales | Gentianaceae | Halenia rhyacophila |
| Lamiales | Orobanchaceae | Castilleja irazuensis |
| Lamiales | Orobanchaceae | Castilleja talamancensis |
| Lamiales | Scrophulariaceae | Buddleja nitida |
| Liliales | Alstroemeriaceae | Bomarea costaricensis |
| Liliales | Alstroemeriaceae | Bomarea hirsuta |
| Malpighiales | Hypericaceae | Hypericum strictum |
| Myrtales | Melastomataceae | Miconia tonduzii |
| Myrtales | Myrtaceae | Ugni myricoides |
| Myrtales | Onagraceae | Fuchsia microphylla |
| Myrtales | Onagraceae | Fuchsia splendens |
| Poales | Bromeliaceae | Puya dasylirioides |
| Poales | Bromeliaceae | Vriesea ororiensis |
| Rosales | Rosaceae | Hesperomeles obtusifolia |
| Rosales | Rosaceae | Rubus eriocarpus |
| Santalales | Loranthaceae | Gaiadendron punctatum |
b. Interacciones planta-insecto
- Se encontró que el principal polinizador durante la estación seca fue Apis mellifera (Hymenoptera, Apidae), la cual está alcanzando su mayor elevación en la zona de estudio. Probablemente se deba a que durante los días despejados, la temperatura elevada durante el día provee de un lugar ideal para estos organismos polinizadores, quienes prefieren recorrer lugares abiertos en lugar de hábitats boscosos.
- Durante la estación lluviosa el polinizador principal es Bombus ephippiatus (Hymenoptera, Apidae), el cual es nativo y se encuentra muy bien adaptado al ecosistema de páramo, aunque cuando hay nubosidad total o lluvia, este desaparece temporalmente.
- No se observó ningún insecto del orden Lepidoptera visitando alguna de las especies de plantas del sitio, a pesar de que muchas plantas de la familia Asteraceae son abundantes en el área de estudio, lo que hace pensar que la limitación es más del tipo climática, que de la estructura funcional de polinizador-planta.
- Durante la estación seca se observaron mariposas, aunque no interactuaron con flores, ya que se desplazaban en vuelos migratorios del Caribe al Pacífico. Posiblemente haya una variedad de mariposas nocturnas, las cuales no fueron detectadas debido a que el estudio se enfocó en los organismos polinizadores diurnos.
Objetivo 2:
a. Organismos polinizadores
- Mediante el método de observación en transectos y captura con redes de niebla, se registraron un total de 543 colibríes pertenecientes a 7 especies.
- En la siguiente figura se presenta la abundancia de las 4 especies que fueron observadas interactuando con plantas en el ecosistema de páramo.
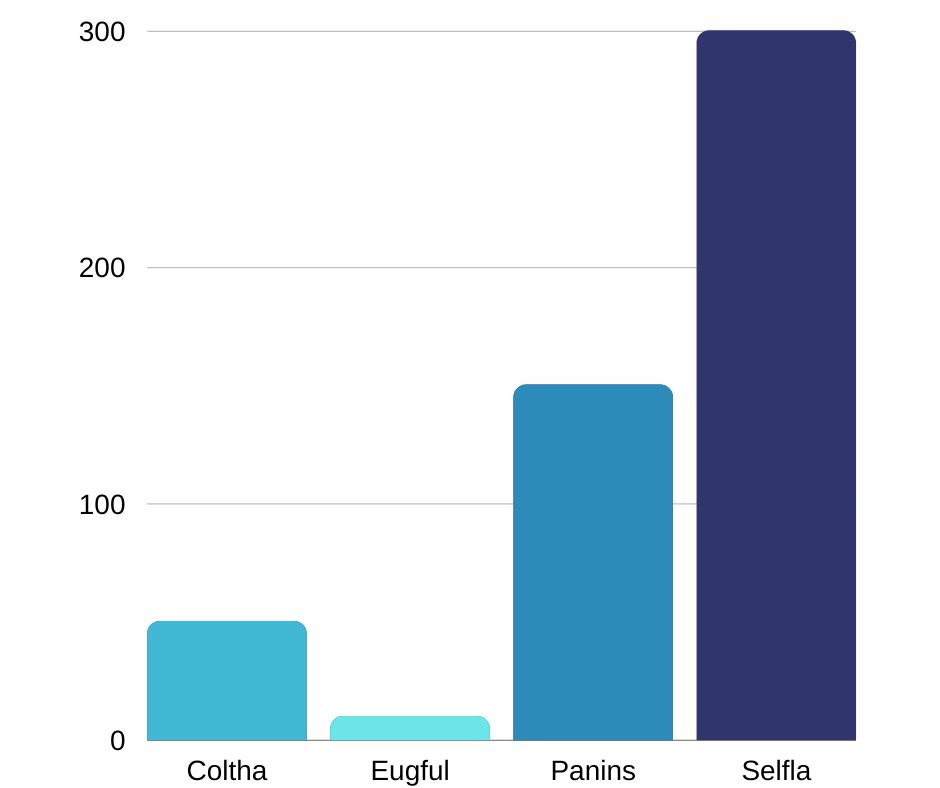
- Durante septiembre y octubre es cuando se registró el mayor número de colibríes, con 89 individuos en cada mes, mientras que en el mes de mayo y julio se registró la menor cantidad de colibríes con 22 individuos en cada mes.
b. Abundancia de flores y fenología floral
- Se analizó la fenología floral durante un año completo en los tres sitios de estudio, cada uno con tres parcelas, de forma que se tiene información de los períodos en que las especies encontradas poseen flores, así como cuando hay mayor floración.
- Las especies de plantas con floración más continua a lo largo del año fueron Hypericum spp. (Hypericaceae) y Castilleja spp. (Orobanchaceae), convirténdose en el alimento más habitual para los organismos polinizadores. En tanto que Hesperomeles obtusifolia (Rosaceae) fue la que atrajo mayor cantidad y diversidad de insectos. Sin embargo, otras especies tuvieron periodos de floración cortos con gran cantidad de flores, tales como Vaccinium consaguineum, Pentacalia firmipes y Pernettya prostrata.
Objetivo 3:
a. Estructura funcional de la comunidad de polinizadores
- Se midieron cuatro rasgos morfológicos en un total de 303 colibríes pertenecientes a las cuatro especies para las que se observó visitación a plantas.
- Selasphorus flammula resultó ser la especie con el pico más corto y menor masa corporal.
- Eugenes fulgens fue la especie con mayor longitud de pico y masa corporal. E. fulgens, como así también P. insignis y C. cyanotus, pueden alimentarse de flores de corola más larga, como efectivamente se observó ya que estas especies se alimentaron frecuentemente de Macleania rupestris y Fuchsia splendens.
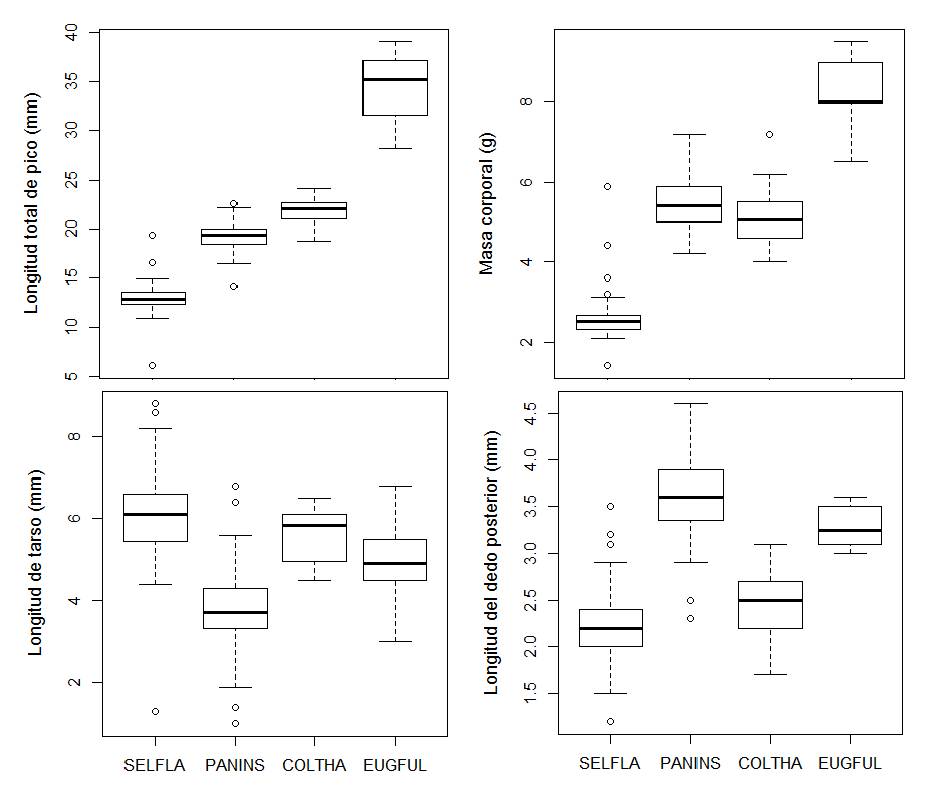
- Se observó una tendencia por parte de S. flammula a alimentarse de flores más pequeñas, por ejemplo, plantas de la familia Asteraceae, mientras que las otras tres especies de mayor tamaño visitaron en general flores más grandes. Lo anterior puede relacionarse con una correlación positiva entre la producción de néctar y el tamaño de las flores (Ornelas et al. 2007) por lo que las especies de mayor tamaño corporal prefieren utilizar flores más grandes que les permita abastecer una mayor demanda energética.
- La gran variación en los valores promedios de los rasgos morfológicos en las diferentes especies de colibríes sugiere una alta diversidad funcional de la comunidad de estos polinizadores en el ecosistema de páramo. Lo anterior puede evidenciar una baja redundancia funcional. Las comunidades ecológicas con baja redundancia funcional pueden tener una baja resiliencia (Pillar et al. 2013), es decir, una reducida capacidad para responder adecuadamente a las perturbaciones ambientales.
Objetivo 4
Analizar la capacidad de las especies de colibríes que habitan el páramo para utilizar nuevos recursos florales.
- Se seleccionaron 2 sitios experimentales para experimentos de trasplante.
- Se seleccionaron 3 especies de plantas con síndrome de polinización de ornitofilia.
- Se tomaron mediciones morfológicas de las especies de plantas que se usarán en experimento de trasplante .
- Se monitoreó la supervivencia de especies de plantas en condiciones de invernadero.

Centropogon valerioi 
Fuchsia splendens
Personas encargadas del proyecto

María Alejandra Maglianesi
(Coordinadora de proyecto) Universidad Estatal a Distancia | Vicerrectoría de Investigación
Escuela de Ciencias Exactas y Naturales
Paul Hanson (Responsable por Universidad)
Universidad de Costa Rica | Escuela de Biología
Alexander Rojas Alvarado (Responsable por Universidad)
Universidad Nacional | Escuela de Ciencias Biológicas
Paola Solera Steller (Responsable por Universidad)
Instituto Tecnológico de Costa Rica | Escuela de Biología
Estudiantes asistentes:
| Estudiantes asistentes: | Institución: | Roles: |
| Alejandro Castro Jiménez | Universidad Estatal a Distancia | Colecta y digitalización datos |
| Angélica Mena Castro | Universidad Estatal a Distancia | Colecta de datos |
| Yahaira Garita Araya | Universidad Estatal a Distancia | Colecta y digitalización datos |
| Marcela Leija Montoya | Universidad Nacional | Colecta y digitalización datos |
| Gerardo Prado Cruz | Universidad Estatal a Distancia | Colecta y digitalización datos |
| Emanuel Brenes Rodríguez | Universidad Estatal a Distancia | Colecta y digitalización datos |
| Douglas Hidalgo Cortés | Universidad Estatal a Distancia | Colecta y digitalización datos |
| Victor Jara Calderón | Universidad de Costa Rica | Colecta de datos y elaboración de mapa (VJ, GH y AV) |
| Gustavo González | Universidad de Costa Rica | Colecta de datos y elaboración de mapa (VJ, GH y AV) |
| Alex Varela | Universidad de Costa Rica | Colecta de datos y elaboración de mapa (VJ, GH y AV) |
| Pablo Muñoz Cambronero | Universidad Nacional | Colecta y digitalización datos de campo |
| Mario Salazar Araya | Universidad Nacional | Colecta y digitalización datos de campo |
| Denis Oviedo Hernández | Instituto Tecnológico de Costa Rica | Colecta y digitalización datos de campo |
| Jessica Madrigal Villalobos | Instituto Tecnológico de Costa Rica | Colecta y digitalización datos de campo |
| Shirley Godínez Mora | Instituto Tecnológico de Costa Rica | Colecta y digitalización datos de campo |
| Otros colaboradores: | ||
| Luis González Arce | Museo Nacional de Costa Rica | Identificación especies plantas |
| Alexander Rodríguez | Museo Nacional de Costa Rica | Identificación especies plantas |
| Luis Alonso Quesada | Museo Nacional de Costa Rica | Identificación especies plantas |
| Alejandra Ospina Pedraza | Centro Agronómico Tropical de Investigación y Enseñanza | Análisis filogenético de plantas |
Universidades involucradas




